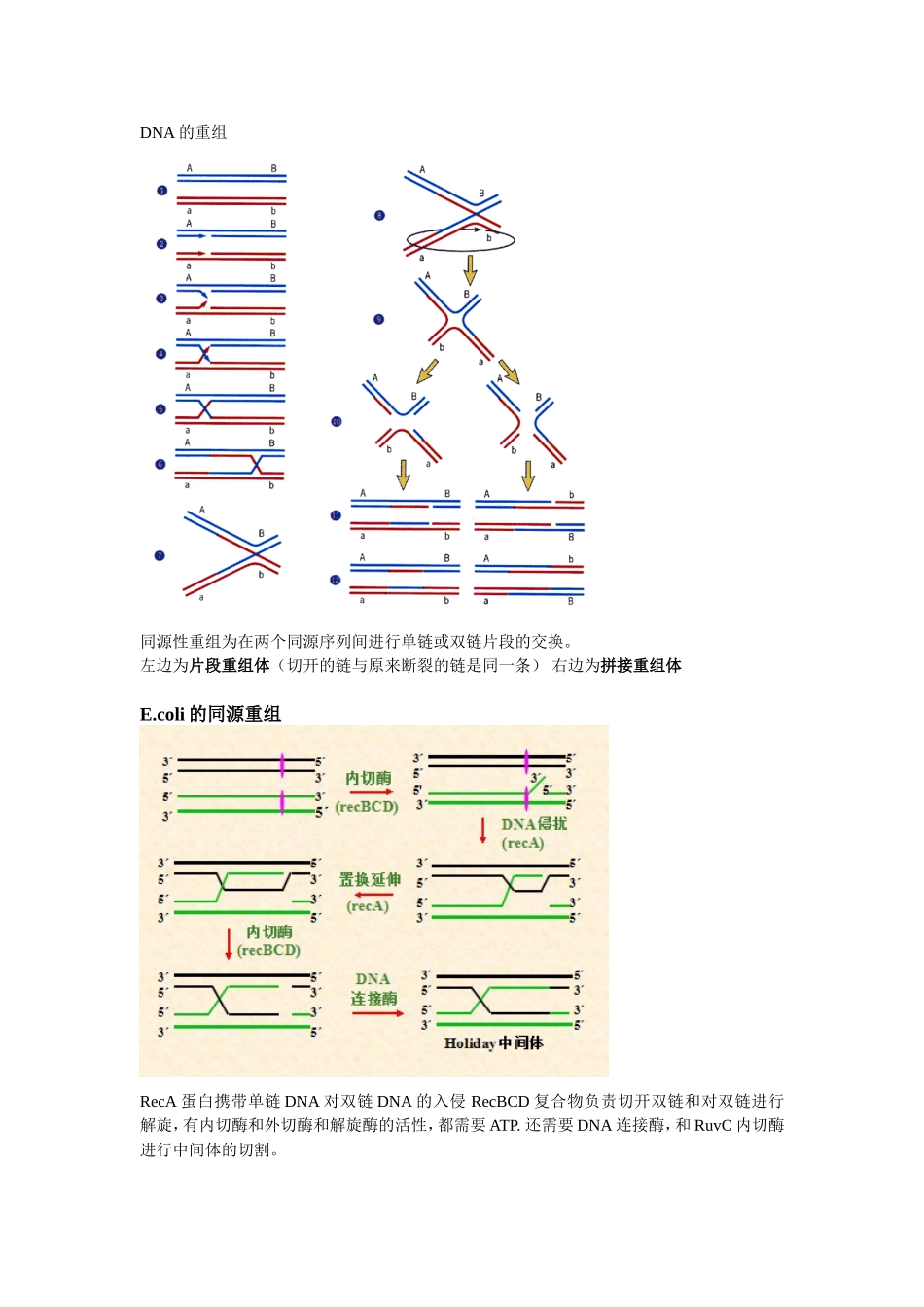
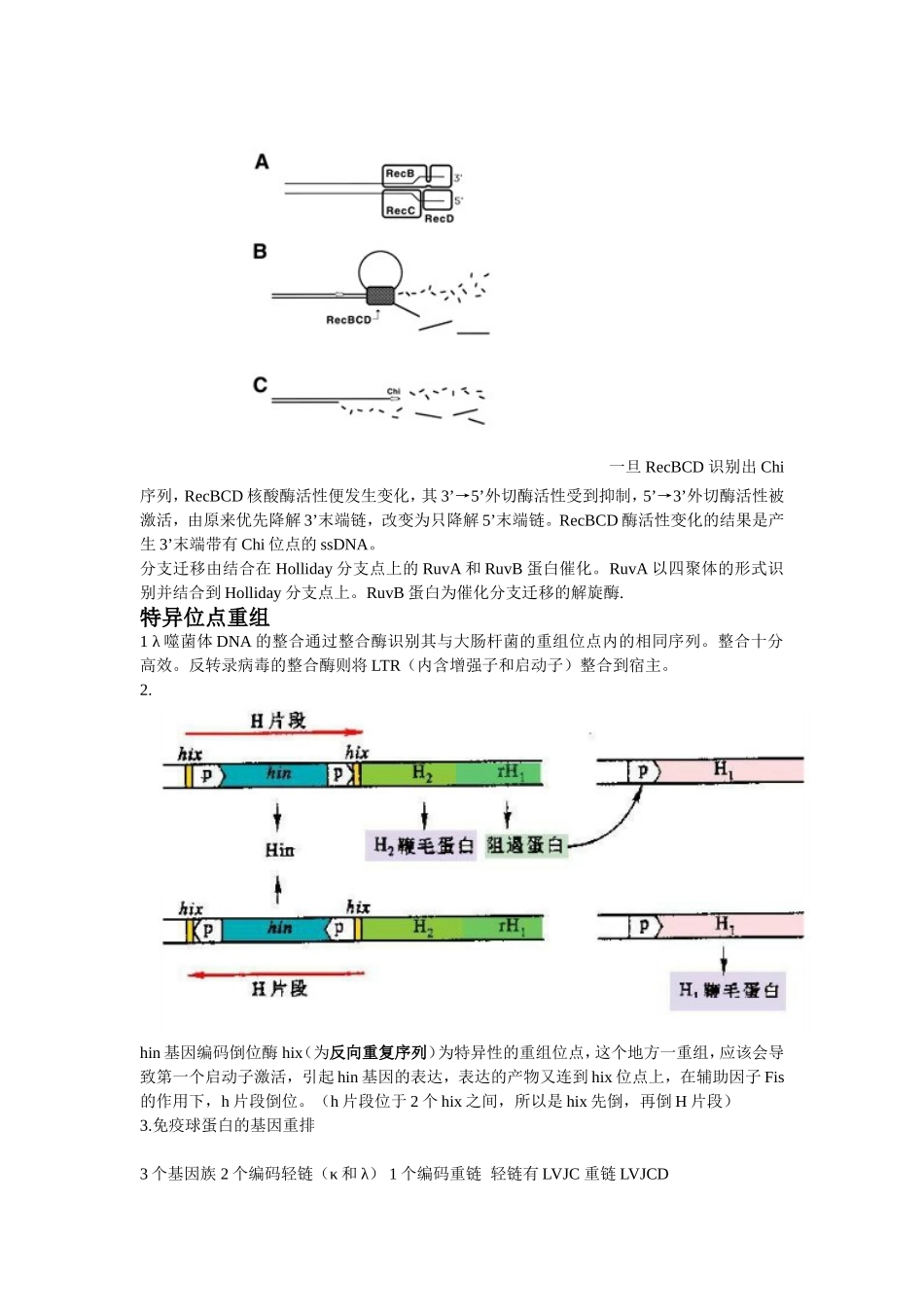
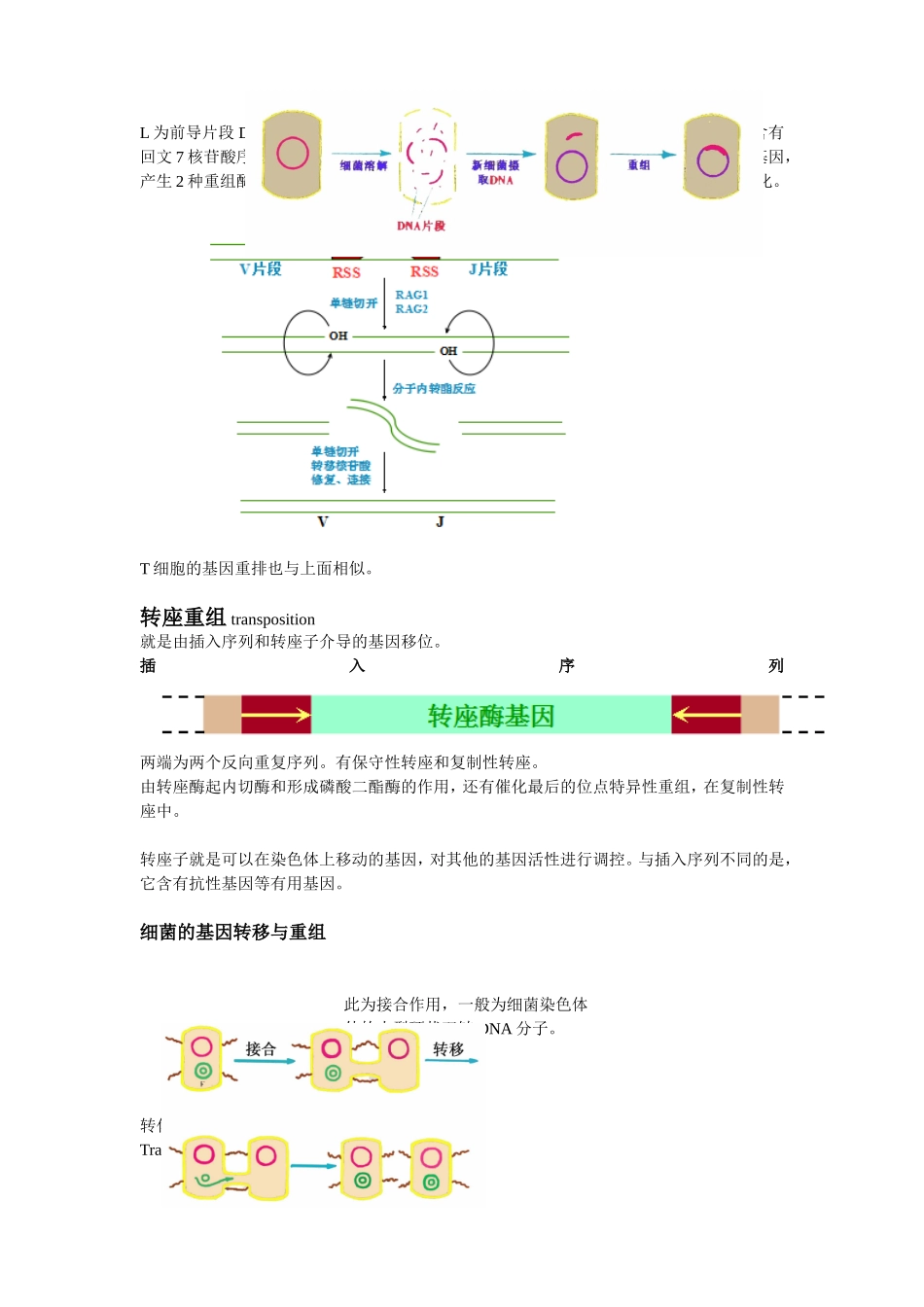
DNA的重组同源性重组为在两个同源序列间进行单链或双链片段的交换。左边为片段重组体(切开的链与原来断裂的链是同一条)右边为拼接重组体E.coli的同源重组RecA蛋白携带单链DNA对双链DNA的入侵RecBCD复合物负责切开双链和对双链进行解旋,有内切酶和外切酶和解旋酶的活性,都需要ATP.还需要DNA连接酶,和RuvC内切酶进行中间体的切割。一旦RecBCD识别出Chi序列,RecBCD核酸酶活性便发生变化,其3’→5’外切酶活性受到抑制,5’→3’外切酶活性被激活,由原来优先降解3’末端链,改变为只降解5’末端链。RecBCD酶活性变化的结果是产生3’末端带有Chi位点的ssDNA。分支迁移由结合在Holliday分支点上的RuvA和RuvB蛋白催化。RuvA以四聚体的形式识别并结合到Holliday分支点上。RuvB蛋白为催化分支迁移的解旋酶.特异位点重组1λ噬菌体DNA的整合通过整合酶识别其与大肠杆菌的重组位点内的相同序列。整合十分高效。反转录病毒的整合酶则将LTR(内含增强子和启动子)整合到宿主。2.hin基因编码倒位酶hix(为反向重复序列)为特异性的重组位点,这个地方一重组,应该会导致第一个启动子激活,引起hin基因的表达,表达的产物又连到hix位点上,在辅助因子Fis的作用下,h片段倒位。(h片段位于2个hix之间,所以是hix先倒,再倒H片段)3.免疫球蛋白的基因重排3个基因族2个编码轻链(κ和λ)1个编码重链轻链有LVJC重链LVJCDL为前导片段D为多样性片段VJ和VDJ重排都是在特异位点,RSS为重组信号序列,含有回文7核苷酸序列,为切割位点,含富含A的九核苷酸序列为识别位点。RAG为重组酶基因,产生2种重组酶。第一次重组在转录前,第二次为受抗原刺激,改变恒定区,发生类别转化。T细胞的基因重排也与上面相似。转座重组transposition就是由插入序列和转座子介导的基因移位。插入序列两端为两个反向重复序列。有保守性转座和复制性转座。由转座酶起内切酶和形成磷酸二酯酶的作用,还有催化最后的位点特异性重组,在复制性转座中。转座子就是可以在染色体上移动的基因,对其他的基因活性进行调控。与插入序列不同的是,它含有抗性基因等有用基因。细菌的基因转移与重组此为接合作用,一般为细菌染色体外的小型环状双链DNA分子。转化作用Transformation转导作用Transduction就是病毒为媒介进行DNA的传播。DNA克隆(DNA重组)将遗传物质与载体DNA结合,形成有自我复制能力的复制子,然后通过转化或转染宿主细胞,再筛选出含目的基因的细胞,对其进行扩增,以获得大量的DNA分子。工具酶限制性核酸内切酶常用Ⅱ类,Ⅰ、Ⅲ类的特异性不高。识别的核苷酸序列为回文序列。切割后产生平端切口、粘端切口。通常识别4、6、8个碱基序列。命名同尾酶不同位点,相同末端(称配伍末端)。同裂酶不同酶,相同位点。碱性磷酸酶能去除核酸分子末端的磷酸基团,防止载体的自连接。有BAP(来自细菌的碱性磷酸酶)和CIP(来自牛小肠的碱性磷酸酶)用完之后要用多核苷酸激酶使其重新磷酸化。TaqDNA聚合酶具有5′→3′聚合酶活性及5′→3′外切酶活性,但没有3′→5′外切酶活性,因而无3′→5′校对活性。因此在PCR中如果发生错配,则没有校正功能。可考虑用新的耐高温酶进行替代。P5′OH3′OH3′P5′图为TA克隆利用Taq酶同时具有末端转移酶的活性,在合成的DNA的3’端加上一个A就可与载体3’端的T连接。cDNA文库和基因组文库都是把基因接种在菌种上,可采用菌落或噬斑原位杂交法进行筛选,探针用目的基因的部分序列或者高度同源基因的部分序列或者推演出来的核苷酸序列。PCR必须已知待扩增基因或DNA两端的序列,并提供合适引物。载体克隆载体和表达载体载体的选择要分子量小,才能容纳足够大的DNA分子,还有数量要够多,2个以上的标记位点,多为抗性基因,β-半乳糖苷酶基因lacZ。还有有多克隆位点(即酶切位点)常用有质粒DNA(位于细菌染色体外)、噬菌体DNA、病毒DNA。1.原核表达体系(E.coli表达体系最为常用)需要一个强的启动子还要有翻译调控序列还要有多接头克隆位点还有有marker当然缺点就是不宜表达真核生物的基因,因其没有进一步的修饰功能,表达的蛋白容易形成不溶性包涵体,很难表达大量可溶性的蛋白。还需要S...